مقدمه
پرندگان برای رشد، تخمگذاری، ایمنی، فعالیت آنزیمی، بازسازی بافتها و غیره به پروتئین نیاز دارند. استفاده از شکل خالص اسیدهای آمینه شاخه دار ایزولوسین و والین در خوراک منجر به کاهش سطح نیاز به پروتئین می شود که این اصل در مورد اغلب اسیدهای آمینه ضروری صادق است. این در حالی است که در خوراک حاوی مقادیر بالای ذرت، همواره با بیش بود لوسین به عنوان دیگر اسید آمینه شاخه دار مواجه هستیم. کاهش سطح پروتئین خوراک پرندگان به دنبال استفاده از شکل خالص اسیدهای آمینه، اغلب منجر به کاهش مصرف آب می شود که دلیل آن کم شدن نیاز به دفع پروتئین مازاد است. این تغییر در ادامه می تواند معضل خیسی بستر و ناهنجاری های پا را در مزارع پرورش و نگهداری طیور صنعتی کاهش دهد. با این حال کاهش سطح پروتئین خوراک به جز جایگزینی اسیدهای آمینه ضروری خالص، به واسطه تنظیم مقادیر این اسیدهای آمینه بر پایه کمینه نیاز مقدور می شود. در این میان با توجه به کاهش ورود اسیدهای آمینه غیرضروری به بدن به موازات کاهش سطح پروتئین علیرغم امکان ساخت این اسیدهای آمینه توسط بدن، بروز کمبود آنها در شرایط ویژه جهت دستیابی به بیشینه ظرفیت فیزیولوژیک بدن محتمل است.
علاوه بر سه اسید آمینه شاخه دار مشهور که در ساختمان پروتئین ها شرکت می کنند، ۲-آمینو ایزوبوتیریک اسید عضو غیرپروتئین زای این خانواده محسوب می شود که تنها در برخی قارچ ها شناسایی شده است. سه اسید آمینه شاخه دار که جزو انواع ضروری طبقه بندی می شوند و بدن حیوانات قادر به سنتز آنها نیست، همچون اسیدهای چرب شاخه دار جزو ترکیبات آلیفاتیک (هیدروکربن غیرآروماتیک) محسوب می شوند. این اسیدهای آمینه دارای یک زنجیره جانبی آبگریز (هیدروفوبیک) متشکل از یک اتم کربن مرکزی متصل به سه یا تعداد بیشتری اتم کربن هستند. این زنجیره جانبی کوتاه و آبگریز هنگامی که وارد ساختار پروتئین می شود، در تعیین ویژگی های دینامیک ملکول از نقش تعیین کننده برخوردار است. ساختار ملکولی لوسین، ایزولوسین و والین به ترتیب ۲-آمینو-۴-متیل-پنتانوئیک اسید، ۲-آمینو-۳-متیل-پنتانوئیک اسید و ۲-آمینو-۳-متیل-بوتانوئیک اسید است. با وجود آنکه اسیدهای آمینه شاخه دار توسط بدن حیوانات سنتز نمی شود اما در حدود ۳۵ تا ۴۰ درصد از نیاز پستانداران و پرندگان به اسیدهای آمینه ضروری، متعلق به این گروه است. نسبت فراوانی این سه اسید آمینه در بدن اغلب حیوانات تقریبا ثابت و معادل ۱.۶:۲.۲:۱.۰ به ترتیب برای والین، لوسین و ایزولوسین است که نشانگر طبیعت مشترک آنها در سنتز و اکسیداسیون است.
برخلاف اسیدهای آمینه شاخه دار دیگر، والین در خوراک طیور بر پایه ذرت و کنجاله سویا محدود کننده است و از طرفی در مقایسه با ایزولوسین، آنتاگونیستی به مراتب بیشتری با لوسین مازاد در جیره غذایی دارد. به همین دلیل در جیره های جوجه گوشتی که با کمینه پروتئین تنظیم می شوند، اغلب نیاز به استفاده از والین خالص سنتتیک وجود دارد. به دلیل ساختار ملکولی مشابه این سه اسید آمینه، مسیر تجزیه آنها تقریبا مشابه است. آنزیم آمینوترانسفراز زنجیره شاخه دار (BCAT) به عنوان اولین آنزیم عمل کرده و پس از آن کمپلکس آنزیمی آلفا کتو اسید دهیدروژناز زنجیره شاخه دار (BCKD) منجر به کاتابولیسم برگشت ناپذیر به اجزای حاوی کوآنزیم آ می شود. تحریک و فعال شدن این مسیر کاتابولیسم توسط یک اسید آمینه شاخه دار که اغلب لوسین است منجر به کاتابولیسم دیگر اسیدهای آمینه شاخه دار نیز می شود. به همین دلیل بیش بود تنها یک اسید آمینه شاخه دار می تواند باعث تخریب و بروز کمبود دیگر انواع آن شود. مطالعات نشان داد که بیش بود لوسین و به میزان کمتر ایزولوسین موجب کاهش سرعت رشد جوجه های گوشتی و زیست فراهمی والین شد. این در حالی بود که عوارض بیش بود والین بر زیست فراهمی لوسین و ایزولوسین به مراتب کمتر و قابل اغماض بود. برخلاف اغلب اسیدهای آمینه که در کبد اکسید می شوند، بیشترین میزان اکسیداسیون اسیدهای آمینه شاخه دار در عضلات و دیگر بافت های محیطی اتفاق می افتد.
بخش بزرگی از اقلام مورد استفاده در تغذیه طیور صنعتی فاقد نسبت مطلوب از اسیدهای آمینه شاخه دار بوده و اغلب سطح لوسین در آنها بسیار بالا است و این مسئله در جیره هایی با پروتئین پایین به دلیل افزایش نسبت غلات که سطح بالاتری از لوسین دارند، شکل جدی تر پیدا می کند. اثرات آنتاگونیست اسیدهای آمینه شاخه دار در چنین شرایطی به ویژه اگر با عدم توازن دیگر اسیدهای آمینه همراه شود، به طور معنی دار سبب اخلال در روند رشد عضلات می شود. به نظر می رسد اثر آنتاگونیست لوسین در جیره هایی با پروتئین کم به مراتب شدیدتر بوده و با افزایش سطح پروتئین، این تاثیر منفی کاهش می یابد.
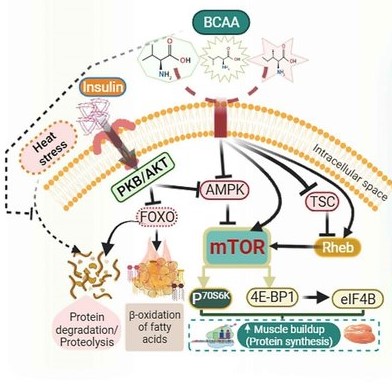
تحقیقات نشان داد کمبود هر سه اسید آمینه شاخه دار به میزان ۵۰ درصد موجب کاهش وزن بدن جوجه های گوشتی در ۲۴ روزگی به میزان یک چهارم گروه شاهد شد. این در حالی بود که کاهش وزن بدن متعاقب دریافت جیره با کمبود ۵۰ درصد اسیدهای آمینه گوگرددار در حدود یک سوم تیمار شاهد بود. از سویی این شدت از کمبود اسیدهای آمینه شاخه دار موجب کاهش شدید وزن نسبی غدد تیموس و بورس فابریسیوس در جوجه های گوشتی شد که حاکی از نقش این گروه از اسیدهای آمینه در توسعه دستگاه ایمنی است. علاوه بر این، نقش اسیدهای آمینه شاخه دار در تنظیم بیان برخی ژن ها مرتبط با سنتز پروتئین در بدن گزارش شده است. اسیدهای آمینه شاخه دار در مجموع یک سوم پروتئین عضلات را به خود اختصاص می دهند و از نقش بسزایی در توقف فرآیند پروتئولیز در زمان تنش گرمایی برخوردار هستند. مطالعات انجام شده روی موش نشان داد که این گروه از اسیدهای آمینه منجر به سرکوب مسیر پروتئولیز لیزوزومال عضلات در کوتاه مدت شده و در بلند مدت نیز پروتئولیز وابسته به ATP در عضلات را کاهش می دهد. در کنار نقش تنظیمی اسیدهای آمینه شاخه دار در تنظیم ساخت و تجزیه پروتئین، کمبود آنها باعث افزایش بتا اکسیداسیون اسیدهای چرب طی مسیر AMPK-mTOR-FoxO می شود. به نظر می رسد لوسین در مقایسه با دو اسید آمینه شاخه دار دیگر، از ظرفیت بالاتری در تحریک سنتز پروتئین از طریق مسیر ام تر (mTOR) داشته و نقش پررنگ تری در تقویت عملکرد ایمنی بدن دارد. ام تر در واقع یک آنزیم تنظیم گر از خانواده پروتئین کینازهای وابسته به فسفاتیدیل اینوزیتول ۳-کیناز است که در تنظیم بسیاری از فرآیندهای سلولی، از نقش تجمیع گر سیگنال های بالادستی از جمله انسولین، انواع فاکتورهای رشد، اسیدهای آمینه و حتی سطح اکسیژن و تعادل انرژتیک برخوردار است. علاوه بر نقش تقویت کنندگی ایمنی، اسیدهای آمینه شاخه دار با تسهیل مسیرهای حذف مشتقات اکسیژن فعال (ROS) منجر به کاهش تنش اکسیداتیو در بدن می شوند.
علاوه بر سه اسید آمینه محدود کننده ذرت و کنجاله سویا (متیونین، لیزین و ترئونین)، اسیدهای آمینه شاخه دار به عنوان عامل محدود کننده بعدی تعریف شده است. این در حالی است که در خصوص اولویت والین و ایزولوسین در شدت محدود کنندگی اختلاف وجود دارد و اختلافات در روش جیره نویسی بر این ابهام افزوده است.
نیاز و نسبت اسیدهای آمینه شاخه دار برای تولید گوشت
تاکنون مطالعات فراوانی در خصوص تاثیر غلظت و نسبت اسیدهای آمینه شاخه دار بر عملکرد رشد و میزان تولید جوجه های گوشتی انجام شده است که اشاره به مهم ترین آنها در این نوشته مدنظر است. بر اساس این یافته ها و دیگر مطالعات، شرکت های تولید کننده سویه های تجاری طیور اقدام به تنظیم و به روز رسانی نیاز اسیدهای آمینه ضروری از جمله انواع شاخه دار می کنند. نیاز اسیدهای آمینه شاخه دار در سویه های صنعتی بر اساس کمینه نیاز پروتئین پرنده تنظیم و پیشنهاد شده است.
طبق یافته یکی از مطالعات جدید، افزایش سطح لوسین از ۱.۸۸ به ۲.۷۳ درصد جیره با حفظ نسبت ایزولوسین و والین به لوسین (به ترتیب ۵۹ و ۶۹ درصد از لوسین) سبب افزایش وزن بدن جوجه های گوشتی به میزان ۴۵ درصد و بهبود مصرف خوراک، ضریب تبدیل غذایی و وزن ماهیچه سینه طی ۲۰ روز دوره انجام مطالعه شد. در مطالعه دیگری مربوط به سال ۲۰۲۰، مکمل سنتتیک ال والین و ال ایزولوسین به جیره حاوی کنجاله بادام زمینی و مخلوط پروتئین حیوانی افزوده شد تا مقادیر این اسیدهای آمینه با جیره های بر پایه ذرت و کنجاله سویا برابر شود (والین و ایزولوسین به ترتیب ۷۶ و ۶۸ درصد لیزین). جیره مبتنی بر کنجاله بادام زمینی و پروتئین حیوانی حاوی مکمل ایزولوسین و والین علیرغم داشتن پروتئین خام کمتر (به میزان ۱.۶۲ درصد) به افزایش وزن و ضریب تبدیل غذایی مشابه با جیره بر پایه ذرت و کنجاله سویا طی ۴۲ روز پرورش جوجه های گوشتی منتهی شد. کمبود ایزولوسین در خوراک (۰.۶۴ درصد) موجب کاهش رشد و افزایش ضریب تبدیل غذایی در جوجه های نر جوان (صفر تا ۱۶ روزگی) شد و این افت عملکرد حتی در شرایطی که سطح دیگر اسیدهای آمینه ضروری خوراک (به جز لیزین و آرژنین) به میزان پنج درصد افزایش یافت، تغییر پیدا نکرد. به نظر می رسد سطح بهینه نسبت ایزولوسین به لیزین (قابل هضم ایلئومی) در جوجه های گوشتی تا ۲۱ روزگی در حدود ۶۶ درصد و پس از آن در حدود ۶۸ درصد است. بدیهی است که پاسخ به ایزولوسین افزوده تحت تاثیر الگوی دیگر اسیدهای آمینه به ویژه لوسین و والین قرار می گیرد.
نتایج مطالعات حاکی از آن است که اثرات مثبت افزودن مکمل ایزولوسین به خوراک تنها در صورتی قابل دستیابی است که نیاز والین تامین شده باشد و بر این اساس میتوان نتیجه گرفت که والین چهارمین اسید آمینه محدود کننده در جوجه های گوشتی است. در میان شاخص های عملکرد، میزان رشد (افزایش وزن) جوجه های گوشتی وابستگی بیشتری به سطح والین در مقایسه با دو اسید آمینه شاخه دار دیگر دارد. این در حالی است که وزن ماهیچه سینه بیشتر تحت تاثیر غلظت ایزولوسین قرار می گیرد و بهبود ضریب تبدیل غذایی و کاهش انباشت چربی در حفره بطنی نیازمند افزایش هم وزن ایزولوسین و والین است. به نظر می رسد که بهترین نتایج عملکرد در جوجه های گوشتی هنگامی به دست می آید که غلظت ایزولوسین خوراک در بازه ۷۰ تا ۸۰ درصد لیزین و سطح والین در حدود ۸۰ درصد لیزین قرار گیرد.
نسبت لوسین به لیزین در جیره های رایج طیور در بازه وسیع تر تغییر می کند. غلظت لوسین معمولا در جیره های پری استارتر بالاتر بوده و در جیره های انتهای دوره به کم ترین مقدار خود می رسد. غلظت لوسین در جیره های جوجه گوشتی به طور میانگین در حدود ۱۳۰ درصد لیزین برآورد می شود. آنچه مسلم است، لوسین به طور اختصاصی سبب تحریک سنتز پروتئین در بدن می شود. در نقطه مقابل مطالعات نشان داد که دریافت لوسین مازاد باعث کاهش قابل توجه والین و ایزولوسین در پلاسمای خون شد. بر این اساس اثرات مثبت لوسین در خوراک جوجه های گوشتی کاملا وابسته به سطوح والین و ایزولوسین یا به عبارتی اقلامی است که در تولید خوراک استفاده می شود. لوسین مازاد در خوراک جوجه گوشتی و در صورت عدم حفظ نسبت آن با دو اسید آمینه شاخه دار دیگر و به ویژه در صورت پایین بودن پروتئین خوراک، قطعا واجد اثرات سمی است.
به طور کلی نتایج مطالعات اخیر حاکی از آن است که افزودن اسیدهای آمینه شاخه دار بدون توجه به نسبت آنها با لیزین نمی تواند از تاثیر قابل توجه روی عملکرد رشد جوجه های گوشتی برخوردار باشد. نتایج آخرین مطالعات بر اساس مدل رگرسیونی در این زمینه حاکی از آن است که نسبت بهینه لیزین، لوسین، ایزولوسین و والین برای افزایش وزن در دوره استارتر جوجه های گوشتی به ترتیب ۱۰۰:۹۴:۵۵:۶۵ است و با نزدیک شدن به انتهای دوره پرورش، این نسبت به ۱۰۰:۱۰۶:۵۶:۷۲ تغییر می یابد. نتایج این تحقیق و مطالعات متاخر حاکی از آن است که نیاز جوجه های گوشتی به اسیدهای آمینه شاخه دار به موازات افزایش سن کاهش می یابد و این حقیقت نافی افزایش نسبت آنها به لیزین در پایان دوره پرورش نیست (با افزایش سن، نیاز به لیزین در مقایسه با اسیدهای آمینه شاخه دار با شدت بیشتر کاهش می یابد).
در جداول زیر تلاش شده تا نیاز اسیدهای آمینه شاخه دار و نسبت آنها با لیزین در آخرین نسخه راهنمای سویه های تجاری متداول جوجه گوشتی ارائه شود:
| سن (روز) | والین | والین/لیزین | ایزولوسین | ایزولوسین/لیزین |
| ۰-۸ | ۰.۸۹ | ۷۳ | ۰.۷۷ | ۶۳ |
| ۹-۱۸ | ۰.۸۵ | ۷۵ | ۰.۷۲ | ۶۴ |
| ۱۹-۲۸ | ۰.۷۶ | ۷۵ | ۰.۶۷ | ۶۵ |
| ۲۹-کشتار | ۰.۷۳ | ۷۵ | ۰.۶۴ | ۶۶ |
| سن (روز) | والین | والین/لیزین | ایزولوسین | ایزولوسین/لیزین |
| ۰-۱۴ | ۰.۹۳ | ۷۴ | ۰.۸۳ | ۶۶ |
| ۱۵-۲۷ | ۰.۸۷ | ۷۵ | ۰.۷۸ | ۶۷ |
| ۲۸-۳۸ | ۰.۸۳ | ۷۷ | ۰.۷۲ | ۶۷ |
| ۳۹-۵۱ | ۰.۷۸ | ۷۸ | ۰.۶۸ | ۶۸ |
| سن (روز) | والین | والین/لیزین | ایزولوسین | ایزولوسین/لیزین | لوسین | لوسین/لیزین |
| ۰-۱۰ | ۰.۹۶ | ۷۵ | ۰.۸۶ | ۶۷ | ۱.۴۱ | ۱۱۰ |
| ۱۱-۲۴ | ۰.۸۷ | ۷۶ | ۰.۷۸ | ۶۸ | ۱.۲۷ | ۱۱۰ |
| ۲۵-۳۹ | ۰.۷۸ | ۷۶ | ۰.۷۰ | ۶۹ | ۱.۱۲ | ۱۱۰ |
| ۴۰-۴۶ | ۰.۷۵ | ۷۸ | ۰.۶۶ | ۶۹ | ۱.۰۶ | ۱۱۰ |

نیاز و نسبت اسیدهای آمینه شاخه دار برای تولید تخم مرغ
اسیدهای آمینه شاخه دار از نقش مهمی در تولید تخم مرغ در مرغان تخم گذار برخوردار هستند و این اهمیت به واسطه نقش تنظیم گر آنها در تولید لیپوپروتئین های زرده از اسیدهای چرب کبد کسب می شود. مقدار نیاز به لوسین در راهنمای تغذیه مرغان تخمگذار سویه لوهمن و هایلاین اعلام نشده است. این در حالی است که در جداول ان.آر.سی (۱۹۹۴)، نیاز به لوسین در مرغان تخمگذار تجاری و مادر برآورد شده است. میزان نیاز به دو اسید آمینه شاخه دار دیگر در راهنمای تغذیه مرغان تخمگذار به تفکیک مقادیر قابل هضم، کل و نسبت به لیزین ارائه شده است.
| سن (روز)/میزان تولید | ایزولوسین | ایزولوسین/لیزین | والین | والین/لیزین |
| ۰-۲۱ | ۰.۷۴ | ۷۰ | ۰.۷۶ | ۷۲ |
| ۲۲-۴۲ | ۰.۷۱ | ۷۲ | ۰.۷۳ | ۷۴ |
| ۴۳-۸۴ | ۰.۶۵ | ۷۴ | ۰.۶۹ | ۷۸ |
| ۸۵-۱۰۵ | ۰.۵۷ | ۷۵ | ۰.۶۱ | ۸۰ |
| ۱۰۶-۱۱۹ | ۰.۶۲ | ۷۹ | ۰.۶۶ | ۸۵ |
| ۱۲۰-۲۴۵ (اولین تخم تا ۲٪، سرانه خوراک ۹۵ گرم) | ۰.۶۷ | ۸۰ | ۰.۷۴ | ۸۸ |
| ۲۴۶-۳۸۵ (پیک، سرانه خوراک ۱۰۰ گرم) | ۰.۶۲ | ۸۱ | ۰.۶۸ | ۸۸ |
| ۳۸۶-۵۰۴ (۸۹-۸۵٪، سرانه خوراک ۱۰۰ گرم) | ۰.۵۹ | ۸۰ | ۰.۶۵ | ۸۸ |
| ۵۰۵-۵۹۵ (۸۴-۸۰٪، سرانه خوراک ۱۰۰ گرم) | ۰.۵۵ | ۷۹ | ۰.۶۱ | ۸۷ |
| پس از ۵۹۶ (کمتر از ۸۰٪، سرانه خوراک ۱۰۰ گرم) | ۰.۵۲ | ۷۹ | ۰.۵۷ | ۸۶ |
| سن (روز)/میزان تولید | ایزولوسین | ایزولوسین/لیزین | والین | والین/لیزین |
| ۰-۲۱ | ۰.۷۱ | ۷۰ | ۰.۷۳ | ۷۲ |
| ۲۲-۴۲ | ۰.۶۶ | ۷۲ | ۰.۶۸ | ۷۴ |
| ۴۳-۸۴ | ۰.۶۱ | ۷۴ | ۰.۶۴ | ۷۸ |
| ۸۵-۱۰۵ | ۰.۴۵ | ۷۵ | ۰.۴۸ | ۸۰ |
| ۱۰۶-۱۱۹ | ۰.۵۶ | ۷۸ | ۰.۶۱ | ۸۵ |
| ۱۲۰-۲۵۹ (اولین تخم تا ۲٪، سرانه خوراک، ۹۸ گرم) | ۰.۶۷ | ۸۰ | ۰.۷۴ | ۸۸ |
| ۲۶۰-۳۳۶ (پیک، سرانه خوراک ۱۱۰ گرم) | ۰.۵۸ | ۷۹ | ۰.۶۴ | ۸۸ |
| ۳۳۷-۴۳۴ (۸۹-۸۵٪، سرانه خوراک ۱۱۰ گرم) | ۰.۵۷ | ۸۰ | ۰.۶۲ | ۸۷ |
| ۴۳۵-۵۳۲ (۸۴-۸۰٪، سرانه خوراک ۱۰۹ گرم) | ۰.۵۶ | ۸۰ | ۰.۶۱ | ۸۷ |
| پس از ۵۳۳ (کمتر از ۸۰٪، سرانه خوراک ۱۰۸ گرم) | ۰.۵۵ | ۸۰ | ۰.۶۰ | ۸۷ |
| سن (روز)/میزان تولید | ایزولوسین | ایزولوسین/لیزین | والین | والین/لیزین |
| ۰-۲۱ | ۰.۷۲ | ۷۱ | ۰.۷۴ | ۷۳ |
| ۲۲-۴۲ | ۰.۶۷ | ۷۱ | ۰.۶۹ | ۷۳ |
| ۴۳-۸۴ | ۰.۶۲ | ۷۴ | ۰.۶۶ | ۷۹ |
| ۸۵-۱۰۵ | ۰.۵۱ | ۷۵ | ۰.۵۵ | ۸۱ |
| ۱۰۶-۱۱۹ | ۰.۵۸ | ۷۸ | ۰.۶۳ | ۸۵ |
| ۱۲۰-۲۵۹ (اولین تخم تا ۲٪، سرانه خوراک ۱۰۳ گرم) | ۰.۶۲ | ۷۸ | ۰.۷۰ | ۸۸ |
| ۲۶۰-۳۳۶ (پیک، سرانه خوراک ۱۰۸ گرم) | ۰.۵۷ | ۷۸ | ۰.۶۴ | ۸۸ |
| ۳۳۷-۴۳۴ (۸۹-۸۵٪، سرانه خوراک ۱۰۹ گرم) | ۰.۵۴ | ۷۷ | ۰.۶۱ | ۸۷ |
| ۴۳۵-۵۳۲ (۸۴-۸۰٪، سرانه خوراک ۱۰۸ گرم) | ۰.۵۳ | ۷۸ | ۰.۵۹ | ۸۷ |
| پس از ۵۳۳ (کمتر از ۸۰٪، سرانه خوراک ۱۰۷ گرم) | ۰.۵۲ | ۷۹ | ۰.۵۸ | ۸۸ |
| سن (روز)/میزان تولید | ایزولوسین | ایزولوسین/لیزین | والین | والین/لیزین |
| ۰-۲۱ | ۰.۷۲ | ۷۱ | ۰.۷۴ | ۷۳ |
| ۲۲-۵۶ | ۰.۶۷ | ۷۱ | ۰.۶۹ | ۷۳ |
| ۵۷-۱۹۹ | ۰.۶۲ | ۷۴ | ۰.۶۶ | ۷۹ |
| ۲۰۰-۱۹۶ (تا ۵٪ تولید) | ۰.۵۱ | ۷۵ | ۰.۵۵ | ۸۱ |
| ۱۹۷-۳۱۵ (سرانه خوراک ۱۰۵ گرم) | ۰.۵۲ | ۷۶ | ۰.۵۶ | ۸۲ |
| ۳۱۶-۴۵۵ (سرانه خوراک ۱۰۵ گرم) | ۰.۵۱ | ۷۸ | ۰.۵۶ | ۸۶ |
| پس از ۴۵۵ (سرانه خوراک ۱۰۵ گرم) | ۰.۴۸ | ۷۸ | ۰.۵۲ | ۸۵ |
طبق گزارش اخیر مکلین و همکاران (۲۰۲۱)، نیاز والین کل و قابل هضم در مرغ تخمگذار به ترتیب ۶۱۴ و ۵۳۲ میلیگرم/روز/پرنده است اما در خصوص ایزولوسین، نتایج مطالعات انجام شده چندان نزدیک نیست. در سال ۱۹۵۵، نیاز لوسین در آمیخته ردآیلندرد و لگهورن سفید تک تاج، کمتر از یک درصد برآورد شد. همچنین برای حفظ تولید تخم مرغ در مرغان لگهورن سفید تک تاج متعاقب دریافت جیره فقیر از نظر ایزولوسین، نیاز به افزودن ۰.۵۳ درصد ال ایزولوسین سنتتیک بود. نتایج مطالعه دیگری در سال ۲۰۲۰ نشان داد که افزودن ال ایزولوسین به جیره با پروتئین کم (دو درصد کمتر از شاهد)، مانع از افت تولید تخم مرغ در فاصله ۲۰ تا ۴۶ هفتگی مرغان تخم گذار شیور سفید شد. در همین مطالعه، مقدار بالای ایزولوسین (۹۰ درصد لیزین) منجر به افزایش تخم مرغ های درشت (۵۶ تا ۶۳ گرم) شد و به همین خاطر، نیاز ایزولوسین ۸۲ تا ۸۸ درصد لیزین قابل هضم ایلئومی برآورد شد. به نظر می رسد افزایش غلظت ایزولوسین در جیره مرغان تخمگذار ضمن افزایش وزن تخم مرغ منجر به کاهش تولید می شود.
ایزولوسین در جیره مرغان تخمگذار از اثر متقابل با والین و لوسین برخوردار است و افزایش آن در جیره های فقیر از نظر والین و لوسین منجر به کاهش سرعت افزایش وزن و توده تخم مرغ شد. مطالعه دیگری نشان داد که بیشترین میزان ابقا نیتروژن در مرغان تخمگذار در دامنه ۰.۴۳ تا ۰.۵۷ درصد ایزولوسین به دست می آید و افزایش غلظت آن به ۰.۸ و یک درصد به ترتیب باعث کاهش وزن بدن و کاهش توده تخم مرغ شد. مطالعه دیگری نشان داد که افزایش غلظت ایزولوسین قابل هضم از ۰.۵۴ تا ۰.۹۴ درصد در جیره هایی با پروتئین کم (۱۴ درصد)، فاقد تاثیر معنی دار بر عملکرد و شاخص های مرتبط با دستگاه گوارش و ایمنی بود. سطح بهینه ایزولوسین، لوسین و والین برای تامین نیاز نگهداری مرغان مادر بر اساس مدل رگرسیونی خطی به ترتیب ۹۴، ۵۲ و ۱۵۵ میلی گرم/کیلوگرم وزن متابولیک/روز (معادل ۳۲۹، ۱۷۲ و ۵۴۶ میلیگرم/کیلوگرم پروتئین خام/روز) برآورد شده است. نتایج یکی از مطالعات اخیر نشان داد که افزایش سطح والین از ۰.۵۹ تا ۰.۷۹ درصد در جیره مرغان تخمگذار در فاصله ۳۳ تا ۴۱ هفتگی (مقدار مصرف خوراک ثابت: ۱۰۰ گرم/روز/پرنده) سبب افزایش توده تخم مرغ، نرخ تخم گذاری، فعالیت تریپسین در دئودنوم و بیان ژن ناقل های اسید آمینه در روده شد. این در حالی است که افزایش والین موجب کاهش واحد هاو، ضخامت پوسته، رنگ زرده تخم مرغ و شاخص های ایمنی شد.
طبق اعلام ان.آر.سی (۱۹۹۴)، نیاز والین و ایزولوسین در فاصله ۲۸ تا ۳۴ هفتگی مرغ تخمگذار به ترتیب ۱۰۱ و ۹۴ درصد لیزین است. برآورد مطالعات جدید حاکی از نیاز کمتر سویه های تجاری مرغ تخم گذار به اسیدهای آمینه شاخه دار در همین بازه سنی بوده و برای والین و ایزولوسین به ترتیب ۹۳ و ۷۹ درصد تعیین شده است.
نیاز اسیدهای آمینه شاخه دار برای دیگر طیور
نیاز اسیدهای آمینه ضروری برای دیگر انواع طیور در برخی منابع مورد اشاره قرار گرفته است اما اطلاعات در خصوص نیاز اسیدهای آمینه شاخه دار اندک است. یکی از معدود بررسی های انجام شده روی اردک در شرایط صنعتی با هدف تولید گوشت نشان داد افزودن سطوح متفاوت ال والین تا سقف ۱.۲۷ درصد به جیره پایه با ۲۹۹۰ کیلوکالری انرژی قابل متابولیسم/کیلوگرم و ۱۸ درصد پروتئین خام فاقد تاثیر قابل توجه بود. مطالعه دیگری که روی بلدرچین های ژاپنی مولد انجام شد نشان داد که بهترین عملکرد (با اختلاف اندک) از نظر رشد و تولید تخم مرغ در پروتئین خام ۱۸ درصد با افزودن ۲۰۰ گرم ال والین به هر تن خوراک به دست آمد. تحقیق بر بوقلمون نشان داد که خوراندن مکمل ال لوسین، ال ایزولوسین و ال والین با نسبت ۳:۱:۲ به جوجه های یک روزه و سپس تغذیه آنها با خوراک عادی سبب افزایش سرعت رشد بدن و وزن ماهیچه سینه شد.
تزریق درون تخم مرغی اسیدهای آمینه شاخه دار
یکی از راهکارهای بهبود عملکرد جنین انواع طیور به ويژه پس از تفریخ، تزریق مواد مغذی یا ترکیباتی همچون هورمون ها به درون تخم است. تزریق ۴۳۵ میلی گرم لوسین و ۲۷۱ میلی گرم ایزولوسین زیر کیسه هوایی تخم مرغ موجب کاهش زمان خروج از تخم مرغ و پیش از تفریخ منجر به تسریع رشد جوجه شد. همچنین تزریق ال لوسین به تخم مرغ سبب بهبود توان تحمل گرما و سرعت رشد جوجه های گوشتی در شرایط تنش گرمایی شد. در مطالعه دیگری مربوط به سال ۲۰۱۷، تزریق محلول ۳۵ میکرومول لوسین در ۵۰۰ میکرولیتر آب مقطر به کیسه زرده تخم مرغ در روز هفتم جنینی سبب کاهش دمای بدن در زمان تفریخ و بهبود وزن جوجه تا پنج روزگی شد. در این مطالعه تزریق والین یا ایزولوسین، چنین تاثیری نداشت. بررسی بعدی همین گروه از محققان نشان داد که تزریق لوسین در روز هفتم جنینی باعث کاهش غلظت تیروکسین و دمای معقدی بلافاصله پس از تفریخ شد. این جوجه ها در روز دهم زندگی و در مواجهه با تنش گرمایی، از دمای معقدی کمتر برخوردار بودند. این نتایج تاییدی است بر نقش لوسین در افزایش تاب آوری گرما در جوجه های گوشتی. همچنین تزریق درون تخم مرغی اسیدهای آمینه شاخه دار به میزان چهار تا هفت میلی گرم در ۰.۵ میلی لیتر آب مقطر به افزایش وزن جوجه یک روزه تا ۲۱ درصد و بهبود شاخص های ایمنی خونی و سلولی منتهی شد. علاوه بر این تزریق ترکیب اسیدهای آمینه شاخه دار (با نسبت ۳:۱:۲ برای لوسین، والین و ایزولوسین) به آمنیون تخم بوقلمون علی رغم کاهش جوجه درآوری به میزان نه درصد، باعث افزایش کیفیت جوجه های تفریخ شده به میزان ۲.۵ درصد و بهبود وزن جوجه ها در زمان تفریخ به میزان ۱۳ درصد شد.
اسیدهای آمینه شاخه دار و سلامت روده، ایمنی و میکروبیوتا
بازدهی و عملکرد رشد و تولید تخم در طیور در گرو توسعه و کارکرد مطلوب دستگاه گوارش، ایمنی و میکروبیوتا است. علی رغم اینکه نقش اسیدهای آمینه شاخه دار در تنظیم و عملکرد طبیعی دستگاه ایمنی و سلامت روده بدیهی است اما تاکنون هیچ ساز و کار مشخصی برای آن کشف نشده است. انتظار بر این است که نقش و ساز و کار احتمالی عملکرد اسیدهای آمینه شاخه دار در طیور مشابه با موش و خوک باشد و با توجه به شمار اندک مطالعات در این عرصه در طیور، می توان به اطلاعات موجود در مورد خوک و موش استناد کرد.
نتایج به دست آمده نشان داد که این اسیدهای آمینه در روده انسان به واسطه ناقل های خنثی و مستقل از سدیم جذب می شوند و انتظار می رود ناقل های مشابه در مغز، عضلات اسکلتی، کبد، طحال و کلیه فعال باشند. همان طور که پیشتر مورد اشاره قرار گرفت، دو آنزیم BCKD و BCAT در ترانس آمیناسیون، دآمیناسیون و اکسیداسیون اسیدهای آمینه شاخه دار فعال هستند و بسته به متابولیت نهایی هر اسید آمینه شاخه دار، لوسین ترکیبی کتوژنیک بوده و والین گلوکوژنیک است. ایزولوسین می تواند گلوکوژنیک یا کتوژنیک باشد. وقتی اسیدهای آمینه شاخه دار تخمیر می شوند، والین به ایزوبوتیرات، لوسین به ۲-متیل-بوتیرات و ایزولوسین به اسید چرب شاخه دار ایزو والرات تبدیل می شود. سلول های ایمنی و مخاطی روده واجد هر دو آنزیم BCAT و BCKD هستند اما غلظت BCAT در کبد اندک است. عضلات اسکلتی که به عنوان بزرگترین ذخایر اسیدهای آمینه شاخه دار محسوب می شوند، موجودی این اسیدهای آمینه را در بدن را از طریق تنظیم شدت اکسیداسیون آنها کنترل می کنند. میزان برداشت اسیدهای آمینه شاخه دار در سلول های ایمنی به طور قابل ملاحظه در مرحله S از چرخه سلولی افزایش می یابد.
نقش اسیدهای آمینه شاخه دار بر ایمنی
والین در میان اسیدهای آمینه شاخه دار کبد، بیشترین توان و سهم تحریک ساخت لنفوسیت گرانولار و آگرانولار و همچنین لنفوسیت های کشنده طبیعی (NK) دارد. اسیدهای آمینه شاخه دار به طور کلی سبب تحریک تولید ایمونوگلوبولین A در غشاهای مخاطی شده و به این ترتیب امکان اتصال عوامل بیماریزا به لامینا پروپیا را کاهش می دهند. اسیدهای آمینه شاخه دار همچنین از مهمترین منابع سوخت برای عملکرد طبیعی گلبول های سفید، تولید سایتوکین های پیش التهابی مانند اینترلوکین ۱۲ و فعالیت سلول های دندریتیک محسوب شده و از نقش ویژه در پاسخ ایمنی ذاتی و تطبیقی بدن برخوردار هستند. از طرفی به نظر می رسد اسیدهای آمینه شاخه دار موجب کاهش نسبت بیان ژن فسفو اف کاپا بی (p-FkB) به ان اف کاپا بی (NFkB) و افزایش نسبت بیان ژن فسفو استات ۳ (p-STAT3) به استات ۳ (STAT3) می شود. ان اف کاپا بی از تنظیم گرهای مهم در پاسخ های ایمنی ذاتی و تطبیقی به حساب می آید که در نتیجه ورود عامل بیماریزا، تنش و سایتوکین های پیش التهابی مانند فاکتور نکروز توموری و اینترلوکین ۱ به سرعت فعال می شود. همچنین اسیدهای آمینه شاخه دار می تواند سبب کاهش اینترلوکین ۶ و فاکتور نکروز توموری آلفا (TNF-ɑ) و از طرفی افزایش ترشح ترکیبات ضدالتهاب شود.
بر اساس مقالات منتشر شده می توان نتیجه گرفت که مقدار و نسبت اسیدهای آمینه شاخه دار علاوه بر نقش آنابولیک، از تاثیر مستقیم بر کارکرد دستگاه ایمنی بدن برخوردار هستند. به طور کلی زیست فراهمی اسیدهای آمینه شاخه دار بر سنتز طبیعی ملکول های سیتوتوکسیک، پلی پپتیدازها، ایمونوگلوبولین ها، سیتوکین ها و همچنین مسیرهای شیمیایی فعال در ایمنی ذاتی و تطبیقی تاثیر غیرقابل کتمان دارد. مقادیر کافی و نسبت مطلوب اسیدهای آمینه شاخه دار همچنین برای ساخت اسیدهای نوکلئیک و پروتئین های اساسی موثر در تکثیر سلول های ایمنی دخالت دارد.
نقش اسیدهای آمینه شاخه دار بر توسعه دستگاه گوارش
در تمام انواع طیور، طول کافی پرزهای (ویلی) روده در راستای ایجاد سطح کافی برای جذب بهینه مواد مغذی ضروری است و به همین دلیل نسبت طول پرز به عمق خمل (کریپت) از شاخص های مهم در برآورد سلامت و کارایی جذب روده باریک طیور محسوب می شود. نقش لوسین در این رابطه بدیهی است و افزودن آن به میزان ۱.۴ تا ۲.۲ درصد خوراک جوجه های گوشتی به طور قابل توجه موجب افزایش طول پرزها در ژژنوم و ایلئوم و بهبود نسبت طول پرز به عمق خمل در هر سه بخش روده باریک شد. همچنین افزودن والین به جیره های کم پروتئین جوجه های گوشتی سبب افزایش طول پرزهای روده و تعداد سلول های جامی (گابلت) در ژژنوم و ایلئوم شد. اسیدهای آمینه شاخه دار طی فرآیند ترانس آمیناسیون می توانند به گلوتامات و آسپارتات تبدیل شوند که سوخت اصلی سلول های مخاطی روده در جذب و انتقال مواد مغذی شناخته می شوند و این احتمال مطرح است که سودمندی اسیدهای آمینه شاخه دار ناشی از اثر مستقیم آنها نباشد.
کمبود اسیدهای آمینه شاخه دار همانند دیگر اسیدهای آمینه به دلیل نقش ساختمانی آنها در سلول های پوششی روده، از نقش اساسی در سلامت روده باریک و توان مبارزه آن با عوامل بیماریزا دارد. علاوه بر این به نظر می رسد اسیدهای آمینه شاخه دار به ویژه ایزولوسین باعث افزایش ترشح بتا-دیفنسین ها از سلول های پوششی روده می شود. دیفنسین ها از پپتیدهای ضدمیکروب به حساب می آیند که برخی از آنها به شکل مستقیم و برخی به واسطه نقش پیام رسانی در دستگاه ایمنی سبب نابودی عوامل بیماریزا شامل قارچ ها، باکتری ها و ویروس ها در سطح سلول های پوششی روده می شوند.
در برخی از مطالعات اخیر، نقش اسیدهای آمینه شاخه دار در افزایش فعالیت تعدادی از ناقل های اسیدهای آمینه در روده مورد اشاره قرار گرفته است که البته نیازمند بررسی های بیشتر و دقیق تر است.
نقش اسیدهای آمینه شاخه دار بر جمعیت میکروبی روده
اطلاعات موجود درباره تاثیر اسیدهای آمینه شاخه دار بر جمعیت میکروبی روده طیور بسیار محدود بوده و نیازمند تحقیقات بیشتری است. بدیهی است اسیدهای آمینه با منشا خوراک علاوه بر میزبان توسط باکتری های روده نیز مورد استفاده قرار می گیرند و در این میان اسیدهای آمینه شاخه دار مستثنی نیستند و به همین سبب بر تنوع جمعیت میکروبی روده موثر هستند. نتایج مطالعات برون تنی (in vitro) نشان داد که اسیدهای آمینه شاخه دار بر جمعیت میکروبی روده و سکوم جوجه های گوشتی موثر هستند. همچنین افزودن اسیدهای آمینه شاخه دار به جیره های فقیر از نظر پروتئین در خوک های جوان موجب افزایش جمعیت و تثبیت باکتری های سودمند در روده باریک شد. به نظر می رسد استرپتوکوکوس لاکتوباسیلوس و کلستریدیوم پرفرنژنس برای رشد و تکثیر نیازمند لوسین و ایزولوسین هستند در حالی که اشرشیاکلی نیازی به این دو اسید آمینه ندارد. بر این اساس شاید بتوان یکی از طرق تاثیرگذاری اسیدهای آمینه شاخه دار بر جمعیت میکروبی دستگاه گوارش را تامین نیاز غذایی انواع خاصی از باکتری ها عنوان کرد. در یکی از مطالعات اخیر، افزودن مکمل والین سبب تغییر الگوی جمعیتی میکروب های روده در مرغان تخم گذار تجاری بالغ شد.
بر اساس تحقیقات انگشت شماری که در این زمینه انجام شده می توان به این نتیجه رسید که تنظیم نسبت و تامین نیاز اسیدهای آمینه شاخه دار در خوراک طیور می تواند از تاثیر سودمند بر جمعیت میکروبی دستگاه گوارش برخوردار باشد و در نهایت منجر به بهبود عملکرد و شاید کاهش نیاز به مصرف آنتی بیوتیک شود.


1 دیدگاه دربارهٔ «نقش اسیدهای آمینه شاخه دار در طیور»
من به عنوان دامپزشک از این مطلب استفاده کردم. تشکر